Gama Casino: Регистрация, Вход, Зеркало, официальный сайт Гама Казино
В период с момента открытия в 2023 г.Gama Casinoпо апрель 2024 г. поднялся в рейтинге похожих сервисов к более старым площадкам. Это произошло благодаря возросшему интересу к онлайн-казино у гемблеров многих стран.
Gama Casino
Владелецказино «Гама»– фирма Fun Fusion N.V. Она работает по адресу ул. Abraham de Veerstraat, д. 9 в столице о. Кюрасао г. Виллемстаде. Остров представляет собой самоуправляемую государственную территорию, где выдают лицензии на деятельность казино более 60 лет.
Торгово-промышленная палата Кюрасао выдала компании Fun Fusion N.V. № N.V. разрешение #365/JAZ Sub-license GLH-OCCHKTW0703052021.
Под ним работаютGama Casinoи другие сервисы, принадлежащие предприятию:
Spark Casino;
Cat Casino;
Daddy Casino.
Проверить подлинность портала можно нажатием на иконку с лицензией в левом нижнем углу экрана.
Для подтверждения безопасности гемблинга игрокам нужно посетить портал ТПП. На нем располагается реестр услуг с поисковой строкой. Надо скопировать в нее название компании и сравнить полученный номер лицензии с тем, который размещен в футере казино.
Основные цвета официального сайта сервиса – темно-серые с яркими слотами для перехода к автоматам и рекламными блоками. В центре располагается краткое описание платформы. Интерфейс пользователя классический, с переводом на русский, немецкий, английский, французский, казахский и узбекский языки.
Страница состоит из таких модулей:
логотипа казино в верхнем углу слева;
полей входа в аккаунт и регистрации в противоположной стороне;
полезных ссылок;
блока быстрого перехода к слотам;
инструкции для игроков;
информационных разделов и правил внизу рабочей области.
Гемблеры, делающие ставки со смартфонов или планшетов, могут перейти по ссылкам для скачивания мобильного приложения для iOS, Android.
Полностью функциональность платформы доступна пользователям после создания аккаунта в сервисе.
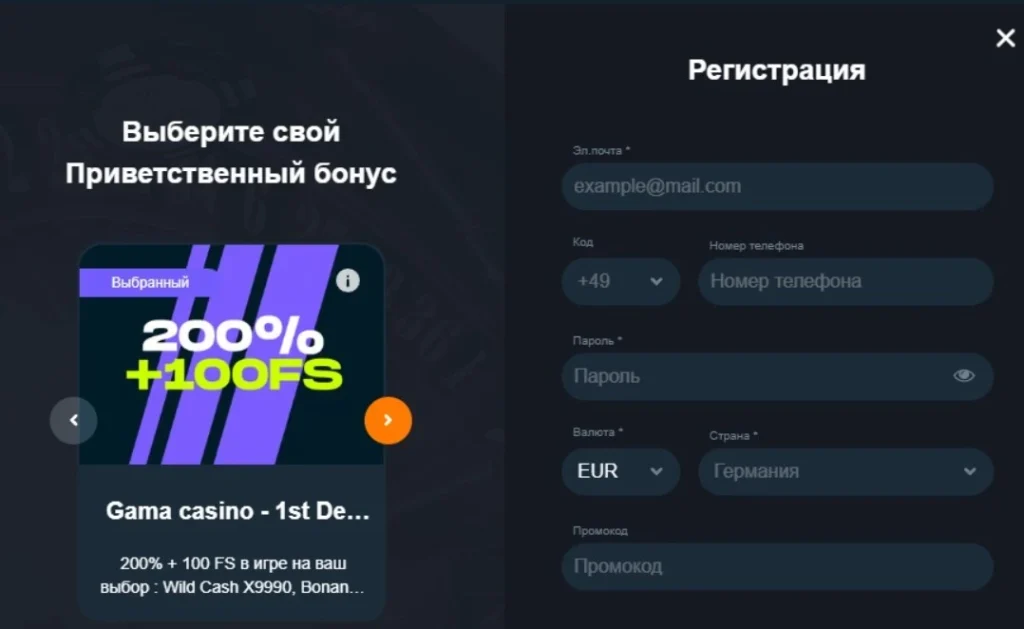
Gama Casino: регистрация личного аккаунта
ВGama Casino регистрация«Личного кабинета» происходит таким образом:
Набрать название онлайн-казино в поисковой строке интернет-обозревателя.
Кликнуть по ссылке для перехода на приветственную страницу портала или рабочего зеркала.
Нажать на поле регистрации.
Заполнить открывшуюся анкету.
Создать пароль для открытия ЛК.
Отметить в выпадающем списке страну проживания.
Выбрать, в какой валюте будет пополняться депозит и выводиться выигрыш.
Подтвердить галочкой свое совершеннолетие.
Принять правила и условия игры в онлайн-казино.
Ввести бонусный код, если он доступен.
Получить СМС или письмо на электронную почту.
Скопировать из него код в пустую строку для подтверждения контактных данных.
Подтвердить свои действия.
Попытаться войти в учетную запись и настроить ее.
В поле анкеты требуется ввод таких данных:
имени и фамилии;
адреса электронной почты;
страны проживания;
типа валюты.
«Гама Casino»: вход в «Личный кабинет»
Для зарегистрированных игроков в«Гама» Casino входв аккаунт представляет собой следующий порядок действий:
Зайти на стартовую страницу оператора.
Кликнуть по кнопке входа.
Набрать логин и пароль от учетной записи.
Подтвердить свои действия или нажать на клавишу Enter.
Gama Casino: зеркало официального сайта
Доступ к порталу затруднен из нескольких государств:
Соединенных Штатов Америки;
Австралии;
Великобритании;
государств Евросоюза;
Грузии и т.д.
Однако гемблеры могут использовать для игры вGama Casino зеркалоплатформы. Так называют полную копию сайта, размещенную в другом домене.
Таким способом пользуются для достижения следующих результатов:
защиты от конкурентов и мошенников;
устранения перегрузок из-за большого количества посетителей;
обхода блокировок провайдера по геолокации;
вывода сервиса из-под поисковых фильтров;
тестирования нового ПО;
входа в казино во время сервисного перерыва.
Получить адрес рабочего зеркала можно такими способами:
от сотрудников службы поддержки в чате или по электронной почте;
в канале мессенджера Telegram;
на сайтах партнеров онлайн-казино;
на профильных форумах.
Получение информации последним способом не самое надежное средство попасть на рабочее зеркало сайта. Из-за того что в онлайн-казино делают ставки на деньги, персональные данные пользователей стали целью хакеров.
Поэтому при входе на такую страницу нужно обратить внимание на следующие нюансы:
цвета оформления портала;
интерфейс;
расположение рекламных модулей;
способы связи со службой поддержки и т.п.
«Гамма Casino»: играть онлайн
В «Гамма Casino» игратьбез регистрации в демонстрационном режиме могут все игроки. Такой способ помогает гемблерам освоить новые слоты, проверить выигрышную тактику. Однако делать ставки на деньги в таком случае нельзя.
Пополнить депозит и вывести выигрыш можно после прохождения верификации.
Процедура такова:
Подготовить сканы документов.
Зайти на приветственную страницу сервиса.
Открыть меню верификации.
Загрузить бумаги.
Дождаться ответа службы поддержки.
Проверка занимает 2–3 дня. В редких случаях процедура затягивается до 2 недель.
Игроки могут подтвердить личность такими документами:
паспортом;
водительским удостоверением;
идентификационной картой;
выпиской из банка;
лично подписанным кредитным договором;
счетом на оплату ЖКХ с адресом регистрации и фамилией.
На платформе установлено программное обеспечение следующих провайдеров:
Endorphina;
Mascot Gaming;
Thunderkick;
Pragmatic Play Live;
Amatic и др.
В верхней части экрана для игроков разместили табло, где они выбирают слоты по нескольким критериям:
новизне игры;
производителю ПО;
популярности;
типу бонуса и т.д.
В разделе слотов чаще всего гемблеры переходят к следующим автоматам:
Hot Fiesta (о музыкальном празднике в мексиканской деревне). В линиях нарисованы национальные музыкальные инструменты – пиньяты, маракасы, барабаны. Лучшие призы выпадают при появлении изображения девушки с розой и мужчины в сомбреро.
Money Train 4 (о путешествии по железной дороге). Главные символы автомата – это пустынная декорация с поездом и барабанами. На них изображены карточные масти и портреты грабителей. Кроме основного выигрыша автомат выдает множители и бесплатные вращения.
Rise of Olympus (о приключениях в Древней Греции). Барабаны здесь представлены игровыми картами, изображениями богов и портретом Титана, заменяющего любую картинку в линиях.
Игроки, желающие сыграть с живыми крупье в атмосфере живой игры, выбирают раздел Live Casino.
Здесь к их услугам представлены развлечения следующих жанров:
рулетки;
блэкджек;
покер;
лотереи;
баккара.
В Live Casino нет демонстрационного режима. Попасть сюда можно после полной регистрации, верификации учетной записи и пополнения депозита.
Gamma Casino: виды бонусов и правила их получения
ПосетителиGamma Casino, впервые регистрирующие аккаунт, получают приветственный бонус.
Его размер зависит от переведенной на баланс суммы и представлен в таблице:
Размер взноса, тыс. руб.
Бонус, %
Количество бесплатных ставок
Условия начисления (отыгрыш вращений)
от 1
100
100
х 40
от 2,5
100
150
х 40
от 10
200
200
х 40
Дополнительные поощрительные программы в сервисе работают таким образом:
Название
Время начисления
Сумма
Участники
Monday Bonus
В 13:00 UTC по понедельникам
5–10% от депозита в зависимости от статуса игрока
Все игроки по программе лояльности
Bonus 35 FS
По средам
35 бесплатных вращений на установленном автомате
Все игроки, выбранные системой в случайном порядке